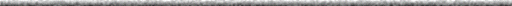|
|
|
| Figure 1. The chain of colored boxes represent the first eight amino acids in the beta chain of hemoglobin. The sixth position in the normal beta chain has glutamic acid, while sickle beta chain has valine. This is the sole difference between the two. |
The molecule, DNA (deoxyribonucleic acid), is the fundamental genetic material that determines the arrangement of the amino acid building blocks in all proteins. Segments of DNA that code for particular proteins are called genes. The gene that controls the production of the beta globin subunit of hemoglobin is located on one of the 46 human chromosomes (chromosome #11). People have twenty-two identical chromosome pairs (the twenty-third pair is the unlike X and Y chromosomes that determine a person's sex). One of each pair is inherited from the father, and one from the mother. Occasionally, a gene is altered in the exchange between parent and offspring. This event, called mutation, occurs extremely rarely. Therefore, the inheritance of sickle cell disease depends totally on the genes of the parents.
If only one of the beta globin genes is the "sickle" gene and the other is normal, the person is a carrier for sickle cell disease. The condition is called sickle cell trait. With a few rare exceptions, people with sickle cell trait are completely normal. If both beta globin genes code for the sickle protein, the person has sickle cell disease. Sickle cell disease is determined at conception, when a person acquires his/her genes from the parents. Sickle cell disease cannot be caught, acquired, or otherwise transmitted. Also, sickle cell trait does not develop into sickle cell disease. Sickle cell trait partially protects people from the deadly consequences of malaria. The frequency of the sickle cell gene reached high levels in Africa and India due to the protection against malaria that occurred for people with sickle cell trait.
|
|
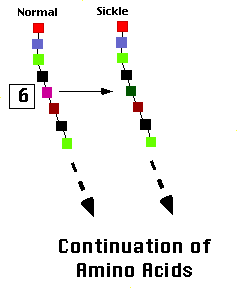
| Figure 2. Normal hemglobin exists as solitary units whether oxygenated or deoxygenated (upper panel). In contrast, sickle hemoglobin molecules adhere when they are deoxygenated, forming sickle hemoglobin polymers (lower panel). |
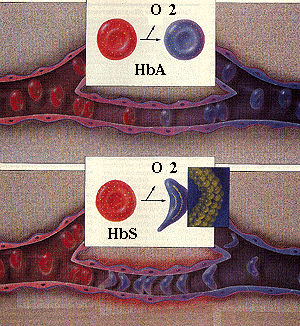
The hemoglobin molecule (made of alpha and beta globin subunits) picks up oxygen in the lungs and releases it when the red cells reach peripheral tissues, such as the muscles. Ordinarily, the hemoglobin molecules exist as single, isolated units in the red cell, whether they have oxygen bound or not. Normal red cells maintain a basic disc shape, whether they are transporting oxygen or not.
The picture is different with sickle hemoglobin (Figure 2). Sickle hemoglobin exists as isolated units in the red cells when they have oxygen bound. When sickle hemoglobin releases oxygen in the peripheral tissues, however, the molecules tend to stick together and form long chains or polymers. These rigid polymers distort the cell and cause it to bend out of shape. While most distorted cells are simply shaped irregularly, a few have a cresent-like appearence under the microscope. These cresent-like or "sickle shaped" red cells gave the disorder its name. When the red cells return to the lungs and pick up oxygen again, the hemoglobin molecules resume their solitary existence (the left of the diagram).
A single red cell may traverse the circulation four times in one minute. Sickle hemoglobin undergoes repeated episodes of polymerization and depolymerization. This cyclic alteration in the state of the molecules damages the hemoglobin and ultimately the red cell itself.
Polymerized sickle hemoglobin does not form single strands. Instead, the molecules group in long bundles of 14 strands each that twist in a regular fashion, much like a braid (Figure 3).

|
| Figure 3. Polymers of deoxygenated sickle hemoglobin molecules. Each hemoglobin molecule is represented as a sphere. The spheres twist in an alpha helical bundle made of 14 sickle hemoglobin chains. |
Despite their imposing appearance, the sickle hemoglobin polymers are held together by very weak forces. The abnormal valine amino acid at position 6 in the beta globin chain interacts weakly with the beta globin chain in an adjacent sickle hemoglobin molecule. The complex twisting, 14-strand structure of the bundles produces multiple interactions and cross-interactions between molecules. The weak nature of the interaction opens one strategy to treat sickle cell disease.
Some types of hemoglobin molecules, such as that found
before birth (fetal hemoglobin),
block the interactions between the deoxygenated hemoglobin S molecules.
All people have fetal hemoglobin in their circulation before birth. Fetal
hemoglobin protects the unborn child and newborns from the effects of sickle
cell hemoglobin. Unfortunately, this hemoglobin disappears within the first
year after birth. One approach to treating sickle cell disease is to rekindle
production of fetal hemoglobin. The drug, hydroxyurea
induces fetal hemoglobin production in some patients with sickle cell disease
and improves the clinical condition
of some people.
|
| Figure 4. Normal red cells maintain their shape as they pass through the capillaries and release oxygen to the peripheral tissues (upper panel). Hemoglobin polymers form in the sickle rell cells with oxygen release, causing them to deform. The deformed cells block the flow of cells and interrupt the delivery of oxygen to the tissues (lower panel). |
The damage to red cell membranes promotes many of the complications of sickle cell disease. Robert Hebbel at the University of Minnesota and colleagues were among the first workers to show that the heme component of hemoglobin tends to be released from the protein with repeated episodes of sickle hemoglobin polymerization. Some of this free heme lodges in the red cell membrane. The iron in the center of the heme molecule promotes formation of very dangerous compounds, called reactive oxygen species. These molecules damage both the lipid and protein components of the red cell membrane. Membrane stiffness is one of the consequences of this injury. Also, the damaged proteins tend to clump together to form abnormal clusters in the red cell membrane. Antibodies develop to these protein clusters, leading to even more red cell destruction (hemolysis).
The anemia in sickle cell disease is caused by red cell destruction, or hemolysis. The production of red cells by the bone marrow increases dramatically, but is unable to keep pace with the destruction. Red cell production increases by five to ten-fold in most patients with sickle cell disease. The average half-life of normal red cells is about 40 days. In patients with sickle cell disease, this value can fall to as low as four days. The volume of "active" bone marrow is much greater than normal in patients with sickle cell disease due to the demand for greater red cell production.
The degree of anemia varies widely between patients. In general,
patients with sickle cell disease have hematocrits that are roughly half
the normal value (e.g., about 25% compared to about 40-45% normally).
Patients with hemoglobin SC disease (where one of the beta globin genes
codes for hemoglobin S and the other for the variant, hemoglobin C) have
higher hematocrits than do those with homozygous Hb SS disease. The hematocrits
of patients with Hb SC disease run in low- to mid-thirties. The hematocrit
is normal for people with sickle cell trait.